Ormar är i regel ganska diskreta, men hasselsnoken tillhör en av de mest anonyma. Man vet relativt lite om den på grund av dess undangömda leverne, och den undgår ofta upptäckt. Om en fara uppstår avslöjar den sig inte, utan ligger kvar tills faran är över eller ringlar lugnt och sakta undan utan att bli upptäckt. Den litar på sitt kamouflage till den grad att även om man t.ex. lyfter på en sten som den ligger under, så ligger den kvar några sekunder för att sedan sakta ringla undan. Om den infångas bits den gärna, men har inga egentliga tänder utan istället några små raspar, vilka sällan går igenom huden. Endast undantagsvis avger den illaluktande analsekret (Andrén & Nilson 1976, Böhme 1993, Larsson 1994).

Arten utnyttjar i hög grad ledningsvärme från sten och mark och anses därmed vara thigmoterm. Detta innebär att den oftast hittas under eller längs med objekt som den kan värma sig mot, och endast mera sällan ses den sola. De individer som solar mera öppet är oftast gravida honor (Spellerberg & Phelps 1977, Luiselli m.fl. 1996). Solbadande sker i regel i skydd av vegetationen (Spellerberg & Phelps 1977).

Hasselsnoken kommer fram på morgonen samtidigt som dagstemperaturen stiger. Den är sedan aktiv några timmar tills kroppstemperaturen nått 29-33°C, då den åter söker skydd för att bibehålla denna temperatur. Den kommer sedan åter fram på eftermiddagen då temperaturen i omgivningen börjar sjunka igen för att behålla den föredragna temperaturen så länge som möjligt. Temperaturkurvan kan beskrivas som en snabb stigning på morgonen för att sedan nå en platåfas kring den optimala temperaturen mitt på dagen, för att sedan långsamt sjunka till dess att den åter söker skydd på kvällen (Spellerberg & Phelps 1977, de Bont m.fl. 1986, Larsson 1994). Vid en undersökning där utlagda plattor använts, tillverkade av plåt och trä, var medeltemperaturen under plattan för hasselsnok 24,2°C (Mutz & Glandt 2004). I samma undersökning tycktes hasselsnok preferera plåt framför trä. Intressant är att i den undersökningen var medeltemperaturen under plattan för kopparödla exakt samma som för hasselsnoken, trots att den föredragna kroppstemperaturen för kopparödla har föreslagits vara så låg som 22,6°C, med intervallet 14,5-28°C (Patterson 1990). Andra undersökningar tyder även på att hasselsnok och kopparödla verkar vara de mest frekvent förekommande arterna under utlagda plattor (Reading 2004).
Studier av hasselsnokar försedda med radiosändare har visat att rörelsemönstret karakteriseras av att arten gärna ligger still långa tidsperioder för att sedan förflytta sig en längre sträcka, som i detta fall är högst 100 meter (Gent & Spellerberg 1993, Larsson 1994).
Det förefaller som om hasselsnoken inte verkar kräva stora habitat på kort sikt, utan uppehåller sig i ett relativt begränsat område. Det bör dock nämnas att i Gent & Spellerberg (1993) hade radiosändaren fästs på kroppen med tejp, vilket fungerade dåligt då sändaren ofta lossnade eller fastnande tillsammans med ormen i vegetationen.
Metoden att operera in en sändare i buken bör ge mer tillförlitliga data än att fästa sändaren på kroppen. I en svensk telemetristudie där sändaren opererats in i buken blev den genomsnittliga förflyttningen under en säsong endast 10,6 meter per dag. Det längsta avstånd någon individ i denna studie befann sig från sitt övervintringsställe var 432 meter. I samma undersökning var det genomsnittliga hemområdet 1,8 ± 0,62 ha (Larsson 1994). Enligt Spellerberg & Phelps (1977) verkar det som att honor rör sig över mindre områden än hanar. Eventuellt gör en del individer längre förflyttningar över längre tidsperioder (Spellerberg & Phelps 1977, Gent & Spellerberg 1993).
Bästa tiden för att träffa på hasselsnok verkar vara i samband med framkomsten på våren, parningen och födseln av ungarna, dvs. april, maj respektive augusti-september (Wolfgang & Meir 1988, Böhme 1993, Larsson 1994, Kéry 2002).
Födoval
Hasselsnoken äter i huvudsak andra reptiler. Den tycks preferera ödlor
av släktet Lacerta i större delen av sitt utbredningsområde
(Andrén & Nilson 1976 Drobenkov 1995, Rugiero m.fl. 1995, Luiselli
m.fl. 1996, Drobenkov 2000, Najbar 2001B), men vad som utgör merparten
av födan kan variera från region till region. Möjligtvis beror
födovalet på vilket bytesdjur som är vanligast i regionen (Rugiero
m.fl. 1995, Reading 2004A), vilket gör arten opportunistisk i sitt födoval.
Huvudfödan i norra delen av utbredningsområdet verkar utgöras
av kopparödla Anguis fragilis (Andrén & Nilson 1976,
1979), men den äter troligen också skogsödla Lacerta vivipara,
huggorm Vipera berus samt insekter i varierande grad (Andrén
& Nilson 1976, 1979). Predation på sandödla Lacerta agilis
av hasselsnok sker troligen i Norden, men arten är alltför sparsamt
förekommande för att kunna utgöra en betydande del av födointaget.
Dessutom finns det studier från England (Spellerberg & Phelps 1977)
som antyder att hasselsnoken inte föredrar sandödla som föda
(baserat på maginnehåll och direkta observationer av födoval).
I populationerna på kontinenten har det påvisats att hasselsnoken
även äter gnagare som fältsork Microtus arvalis, dvärgmus
Micromys minutus, mindre skogsmus Apodemus sylvaticus i varierande,
men ofta sparsam, utsträckning. Även näbbmöss Sorex
äts. Det verkar dock som om det är mest ungar i boet som tas (Drobenkov
1995, Rugiero m.fl. 1995, Luiselli m.fl. 1996, Najbar 2001B). I Norden förefaller
det som om predation på gnagare sker endast undantagsvis (Andrén
& Nilson 1976, Larsson 1994), vilket gör de nordiska populationerna
särpräglade i sitt bytesval. Det har även observerats att hasselsnok
försökt ta boungar hos fåglar, i detta fall sånglärka
Alauda arvensis (Najbar 2001A). Predation på snok Natrix
natrix verkar vara sällsynt och eventuellt lokalt förekommande
(Böhme 1993, Najbar 2001B). Det har även påvisats predation
på aspisvipera Vipera aspis i Medelhavsområdet (Rugiero
m.fl. 1995). Eventuellt skiljer sig födovalet mellan vuxna honor och hanar
såtillvida att honorna äter mer sällan än hanarna och tar
större byten som ormar och gnagare, medan hanarna äter oftare och
tar mest mindre byten som ödlor (Luiselli m.fl. 1996).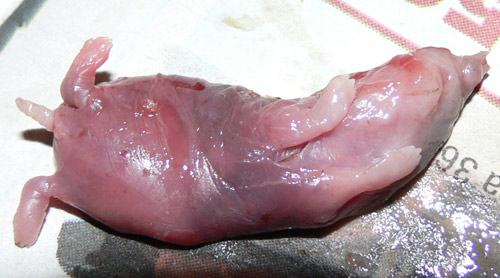

Första bilden föreställer innehållet i magen på en död hasselsnok jag hittade: bounge, troligen åkersork. Andra bilden är ett av favoritbytena: kopparödla.
Populationerna av hasselsnok i England har visat sig leva till största
delen på smågnagare, medan reptiler, då vanligen skogsödla,
äts i störst utsträckning av unga individer (Spellerberg &
Phelps 1977, Goddard 1984). Detta innebär att hasselsnoken i så fall
skulle vara näringskonkurrent till huggormen (predation på huggorm
av hasselsnok har inte påvisats här). Det har även visat sig
att de båda arterna inte heller förekommer i samma områden
i England (Spellerberg & Phelps 1977), vilket även rapporteras från
Vitryssland (Drobenkov 1995).
Förekomsten av insekter i magen på en hasselsnok kan mycket väl
komma från magen på en insektätande ödla som hasselsnoken
har ätit. Insekter som kommer från ödlor är i regel tuggade
(Najbar 2001B). Det har dock observerats att hasselsnokar aktivt äter insekter
(Spellerberg & Phelps 1977). Det har även hittats hela större
bitar av insekter magen på hasselsnok som antyder att den själv har
ätit dem (Najbar 2001B).
Hasselsnoken är även kannibal i varierande grad, vilket troligen är
en del av regleringen av dess ofta individfattiga populationer. I en undersökning
i Vitryssland, Lettland och Ukraina utgjorde kannibalism ca 10 % av födointaget,
baserat på maginnehåll av frilevande hasselsnokar (Drobenkov 1995).
Hasselsnokens tillvägagångssätt för att fånga reptiler
tycks vara att snabbt bita tag kring området på bytets huvud, för
att sedan svälja detta först. Hasselsnoken kan även lägga
ett par slingor av sin kropp kring offrets för att hålla fast det
medan den sväljer. Intressant är att huggormen inte verkar använda
sina tänder till försvar, samt att hasselsnoken är motståndskraftig
mot huggormens gift (Andrén & Nilson 1976).
Reproduktion
Hasselsnokens parningsperiod påbörjas i regel kort efter framkomsten
på våren, i Sverige under andra halvan av april (Andrén och
Nilson 1976). På kontinenten kommer hasselsnoken fram från hibernationen
i slutet av mars eller i början av april (Wolfgang & Meier 1988, Böhme
1993, Najbar 2001A), i England i slutet av april (Spellerberg & Phelps 1977).
Hanarna och de icke könsmogna individerna kommer fram först (Spellerberg
& Phelps 1977, Wolfgang & Meier 1988, Luiselli m.fl. 1996). I de mera
centrala delarna av utbredningsområdet är även höstparning
dokumenterat (Böhme 1993).
Det kallare klimatet i de nordliga delarna av utbredningsområdet gör
att reproduktionscykeln är förlängd till två år, vilket
innebär att reproduktion endast sker vartannat år (Andrén &
Nilsson 1976). I Polen finns det uppgifter som antyder att reproduktion sker delvis
varje år, delvis vartannat år (Najbar 2001A), medan det i Vitryssland
sker reproduktion varje år (Drobenkov 2000). Luiselli m.fl. (1996) anger
att reproduktion i Italien sker vartannat eller till och med vart tredje år.
En längre studie i England (Reading 2004B) tyder på att viktiga faktorer
för att honan ska kunna reproducera sig är honans kondition samt födotillgången
under året. Värt att notera i denna undersökning är att det
inte togs hänsyn till mängden smågnagare i beräkningen av
bytestillgång då de ansågs vara för sparsamt förekommande
för att ha betydelse som föda.
Parningen föregås av hanarnas kamper där de även biter varandra.
Hanarna uppvisar ofta sår på kroppen under våren. Även
under själva parningen biter hanen fast sig i nacken på honan. Detta
beteende att bitas i samband med parningen är ganska ovanligt bland ormar
och brukar benämnas som typiskt ödlebeteende (Andrén & Nilson
1976, även Rüdeberg 1955).
Hasselsnokshonor antas bli könsmogna vid fyra års ålder och ungefär
50 cm längd (Spellerberg & Phelps 1977, Najbar 2001B). I en omfattande
undersökning i England under 10 års tid (Reading 2004A) förökade
sig honorna i regel först vid 6-7 års ålder, vilket kan bero
på konkurrens mellan dem. Den genomsnittliga nos-kloak längden var
här 43,5 cm. Exempel finns dock på andra populationer av hasselsnok
i England där honorna reproducerar sig redan vid 34 cm nos-kloak längd
(Goddard 1984). Även i Italien har det påträffats gravida honor
med en kroppslängd på 43,5 cm (Luiselli m.fl. 1996). Konkurrens mellan
honor gör troligtvis att endast ett fåtal av dem, dvs. de största,
reproducerar sig. Den större kroppsstorleken ger en fördel vid födovalet
då de kan ta större byten (Drobenkov 2000, Reading 2004B). Då
det krävs mycket energi för hasselsnoken att reproducera sig kan endast
stora honor reproducera sig i större utsträckning (Luiselli m.fl. 1996).
Hanen antas bli könsmogen vid tre till fyra års ålder och 45-50
cm kroppslängd (Spellerberg & Phelps 1977, Luiselli m.fl. 1996, Najbar
2001A).
Ungarna föds från början av augusti till slutet av september i
Skandinavien i samband med att ormarna uppsöker övervintringsplatser
(Gislén & Kauri 1959, Andrén & Nilson 1976, 1979), vilket
i grova drag verkar gälla för hela utbredningsområdet (Spellerberg
& Phelps 1977, Wolfgang & Meier 1988, Drobenkov 2000, Najbar 2001A). Tidpunkten
för födseln verkar bero på väderförhållandena.
Om det är en kall vår och försommar sker födseln senare än
om det är varmt (Böhme 1993, Najbar 2001A). Ungarna föds genom
ovovivipari, dvs. ägghinnan går sönder i samband med födseln.
Svenska uppgifter anger kullstorleken till 7-15 individer med en längd på
14-21 cm (Gislén och Kauri 1959, Andrén & Nilson 1976). Liknande
siffror anges även från Polen (Najbar 2001A) och Italien (Luiselli
1996). Honans dräktighetsperiod anges vara tre till fyra månader (Spellerberg
& Phelps 1977, Wolfgang & Meier 1988). Det har även påvisats
en positiv korrelation mellan honans storlek, antalet ungar samt andelen levande
ungar som föds (Luiselli m.fl.1996).
Sällan ses det ett- och tvååringar i naturen, vilket talar för
annorlunda levnadssätt än de vuxna individerna, låg överlevnad
eller helt enkelt låg rekrytering (Spellerberg & Phelps 1977, Goddard
1984, Reading 2004A). Kannibalism skulle eventuellt kunna vara en viktig faktor
för att förklara den låga andelen juvenila individer (Drobenkov
1995). Drobenkov (2000) nämner dessutom att dödligheten för juvenila
individer är mycket hög, över 80 %. Han har dock baserat detta
på frånvaron av unga individer snarare än konstaterad mortalitet.
Även perioden mellan födseln och övervintring för ungarna
bör ha stor betydelse för överlevnaden (Najbar 2001A).